Differences in larval phenotype between eastern and western populations of Sphinx chersis (Lepidoptera: Sphingidae)
Sphinx chersis (Hübner, 1823), the great ash sphinx, has an unusually disjunct geographic distribution throughout North America, with populations largely concentrated in two regions: 1) the northeastern USA and southeastern Canada in the east and 2) the southwestern USA in the west. The species is scarce to absent in the southeastern USA, Great Plains, and Pacific Northwest, separating eastern and western populations by considerable distances. Albeit, its current eastern range has apparently recessed in recent decades due to the destruction of Fraxinus host plants by the invasive emerald ash borer, Agrilus planipennis (Wagner & Todd 2016). Still, even historically the species was probably only weakly contiguous across the Great Plains.
The unusual distribution of S. chersis has naturally provoked the question of whether it represents multiple cryptic, allopatric entities. Populations in the west are sometimes split into subspecies oreodaphne (Edwards, 1874; TL: California) and pallescens (Rothschild & Jordan, 1903; TL: Arizona), and those in Mexico treated as subspecies mexicanus (Rothschild & Jordan, 1903; not treated in this study due to insufficient records). Geographical variation in size and wing shape have been noted, with western adults being smaller in size and having a sharper forewing apex compared to eastern adults (Tuttle 2007; Kesting-Handly & Koiber 2018). Recently, it was also realized based on Arizona specimen that the larval phenotype and DNA barcoding of western populations differs significantly from that of the eastern, providing increasing evidence that the former may represent a separate species (Wagner & Todd 2016).
Given this possibility, it would be greatly beneficial to have a better understanding of the phenotypic differences between eastern and western larvae, prompting the current study. Previous descriptions found in the literature, such as those given in Wagner (2005), Tuttle (2007) and Powell & Opler (2009), only treat one or the other population without comparative context of the other (eastern for the former and western for the latter two, respectively), unbeknownst that they may be describing distinct taxonomic units. Through an extensive examination of larval photographs, I present detailed descriptions and illustrations of the (fifth instar and prepupal) larval phenotype and geographic distribution of eastern and western populations of S. chersis and discuss key diagnostic differences.
MATERIALS AND METHODS
Photographs of S. chersis larvae available on the citizen science platform iNaturalist (https://www.inaturalist.org/observations?place_id=any&taxon_id=127169) were examined. As this platform is prone to misidentifications, I was careful to ignore any larvae that were not S. chersis.
RESULTS AND DISCUSSION
Description of eastern population larva. Fifth instar. The phenotype of the fifth instar larva is perfectly consistent with that described and depicted in Wagner (2005), which is unsurprising given that he based his description on larvae taken from eastern populations. In the green form (Fig. 1), the dorsal abdomen is often whitish green whilst the rest of the body is typically a lime green. There is moderate to heavy speckling of white granules in the thoracic region and below the spiracles. With no exceptions, the spiracles have black centers with white rings (eastern larval records with orange spiracles are almost certainly misidentifications, typically of the similar Ceratomia undulosa). The oblique stripes along the body are usually weakly to moderately well developed, whitish, and unedged with any other colors in the pure green form. The horn is typically bright blue but occasionally pink.
A pink form (Fig. 2) and their intermediates occur at an uncommon but appreciable rate (23/492 = 4.67% of records examined); in the most extreme manifestations, the ground color is bright yellow dorsally and pink below the spiracles, the oblique stripes are heavily edged with pink, and all sclerotized parts (head, horn, etc.) are completely pink. This form enhances crypsis on the unique red autumn foliage of white ash (Fraxinus americana), the most common ash species on the east.
Prepupa. In the prepupal stage (Fig. 3), the larva usually exhibits no color change unlike many sphinx species (records of wandering prepupae that are brownish on the east are usually misidentified C. undulosa, or a pink form S. chersis).
1 
2 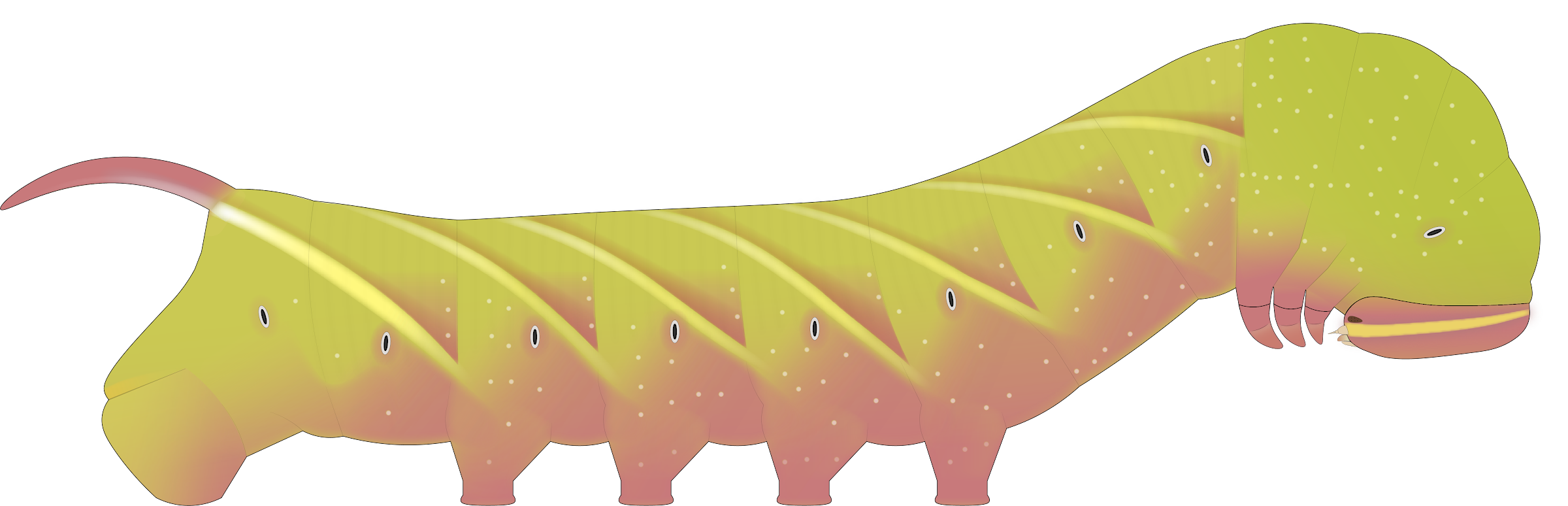
3
FIGS. 1-3. Illustrations of eastern S. chersis larvae. 1, Fifth instar, green form. 2, Fifth instar, pink form. 3, Prepupa, green form (identical to fifth instar).
Description of western population larva. Fifth instar. The larva of western populations (Fig. 4) differs in several significant ways from that of eastern populations. The dorsal abdomen has a bright green cast rather than a whitish cast and the rest of the body is a pale, turquoise green: essentially the reverse ground coloration scheme as eastern larvae. There is much lighter speckling of white granules, largely limited to the thoracic region. Strikingly, larvae on the west generally have spiracles with orange centers rather than black ones, especially in the desert regions (Arizona, New Mexico, and adjacent regions), although exceptions were found. The oblique stripes along the body are usually more strongly developed and solid white, sometimes thinly edged with dark teal or purple, especially in desert larvae.
Records of pink forms are lacking and likely less common than on the east, though large-scale larval rearing may be needed to quantitively verify this. The common ash species on the west, velvet ash (F. velutina) and Oregon ash (F. latifolia), do not have red autumn foliage.
Prepupa. During the prepupal stage (Fig. 5), western larvae display a vivid color change to a deep amber to purplish brown dorsally. This life history trait objectively unifies and distinguished all western larvae from eastern ones.
4 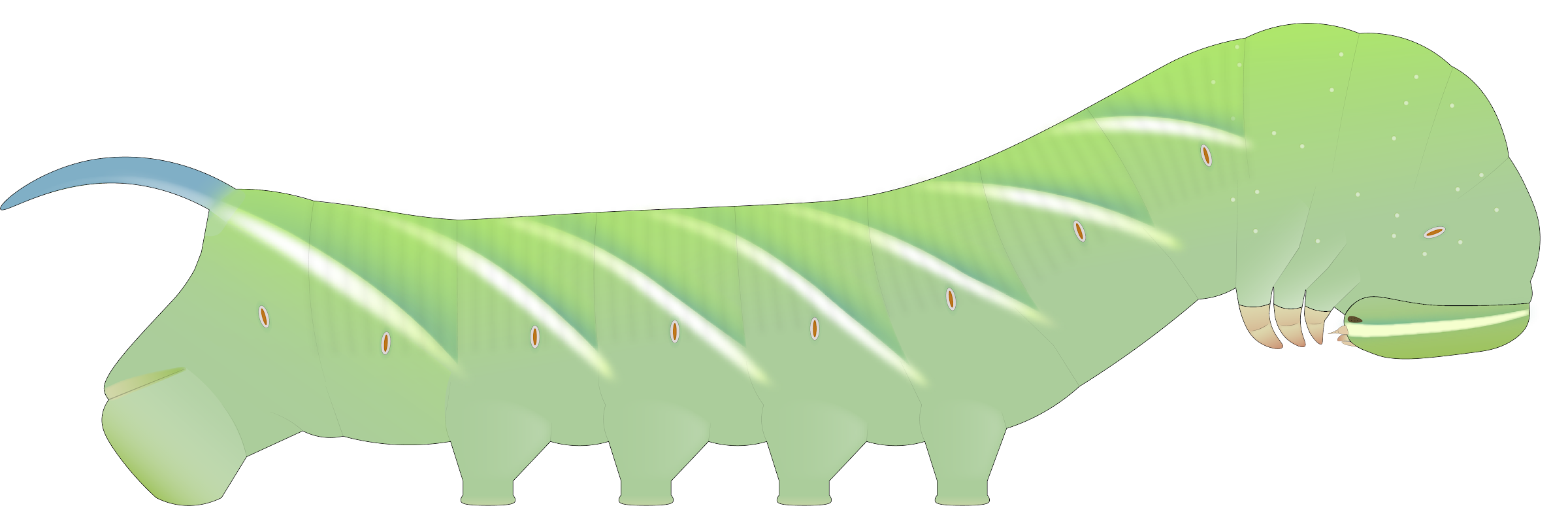
5 
FIGS. 4-5. Illustrations of western S. chersis larvae. 4, Fifth instar, green form. 5, Prepupa, green form, displaying color change.
Distribution. Examined S. chersis larval records with either the eastern or western phenotype formed well defined, allopatric distributions (Fig. 3). Those with the eastern phenotype occurred predominantly throughout New England, the Midwest, and southern Canada. A few surprising records occurred in Montana, Nebraska, Colorado, and New Mexico at the western extremities. Records from the latter two states were the sole exceptions that were in proximity with those with the western phenotype and suggest that the two entities may be sympatric along the eastern foothills of the Rocky Mountains. It is noteworthy, however, that all records from Canada, even far west to Alberta, displayed the eastern phenotype; populations in western Canada are considerably further north than most populations with the western phenotype.
Those with the western phenotype occurred predominantly throughout the desert Southwest and Great Basin, with somewhat spottier records along the northern California coast up to extreme southern Washington. A few Mexican records occurred in the immediate vicinity of the southwestern USA records, which were assumed to belong to the western entity that has been described here rather than the mexicanus subspecies, if distinct. Although larval photographs are missing from the heart of Mexico, based on geography, one would imagine that mexicanus is more closely allied to the western entity than it is to the eastern one.
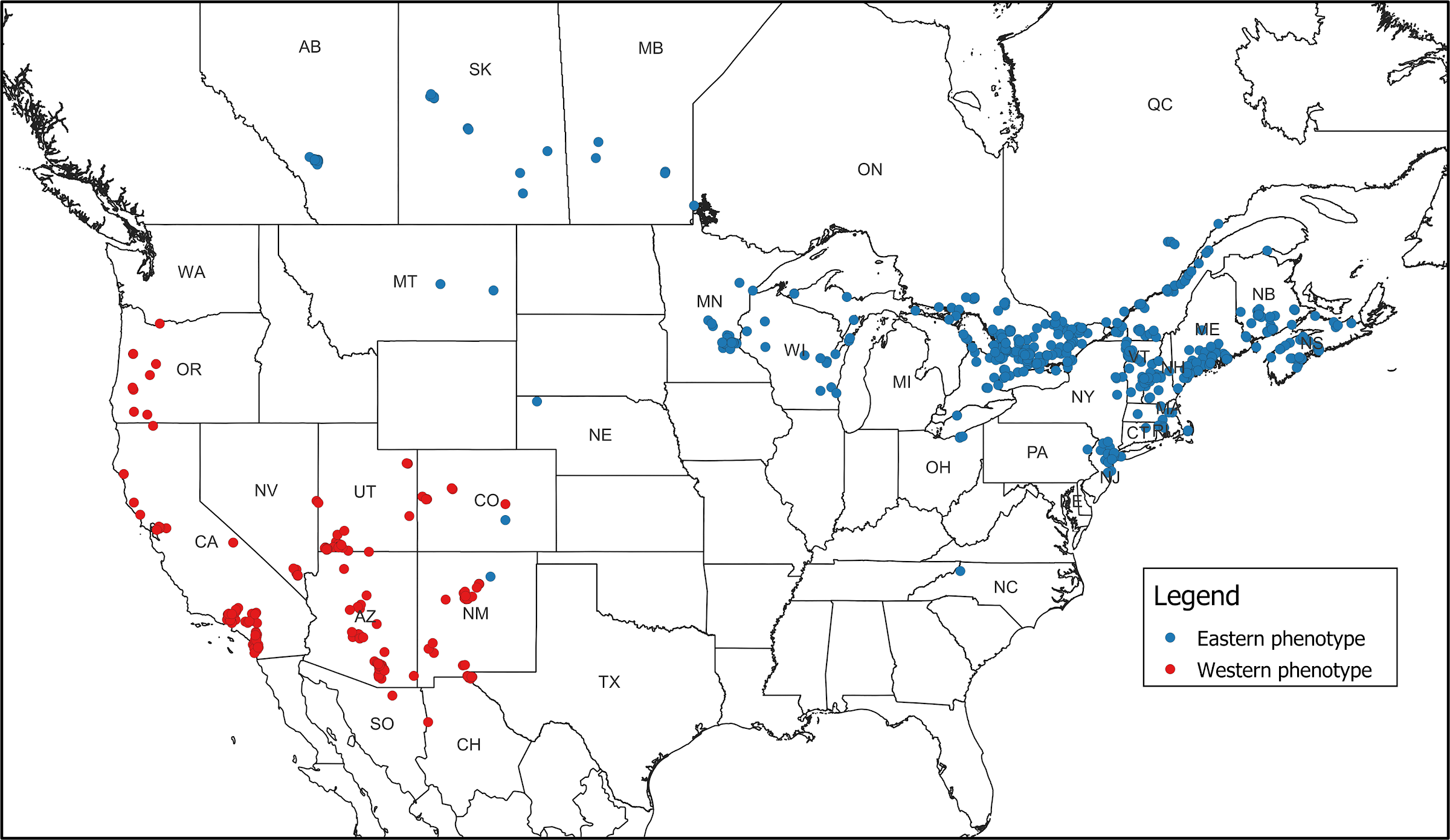
FIG. 3. Distribution of examined larval records of S. chersis from iNaturalist, which displayed either the eastern phenotype (blue) or the western phenotype (red).
Conclusion. Overall, the single most objective and diagnostic difference between the eastern and western populations of S. chersis is that the prepupa becomes deep brown in the west, whereas there is little to no change in the east. There are also notable differences in final instar ground color, spiracle color, speckling, oblique stripes, and occurrence of pink forms. The two entities do not appear to be significantly sympatric in any of their range, except along the eastern foothills of the Rocky Mountains.
The degree of larval and distributional differences seen in eastern and western populations of S. chersis is comparable to that of other eastern/western sphingid sister pairs such as Pachysphinx modesta/occidentalis and Hemaris diffinis/thetis. Likewise, this stark geographic dichotomy in larval phenotype would be highly unusual for a single species to exhibit. Thus, based on larval phenotype and distribution alone, a compelling argument could be made for a taxonomic distinction between eastern and western populations. A more extensive molecular survey of these populations is needed to confirm whether the magnitude of their phenotypic differences is reflected at the molecular level.
LITERATURE CITED
Kesting-Handly T., S. Kloiber. 2018. Sphinx chersis, Sphingidae of the United States. Available from: https://www.sphingidae.us/sphinx-chersis.html (October 28, 2021).
Powell J.A., P.A. Opler. 2009. Moths of Western North America. University of California Press. Berkeley, CA. 243 pp.
Tuttle J.P. 2007. The hawk moths of North America: A natural history study of the Sphingidae of the United States and Canada. The Wedge Entomological Research Foundation. Washington, D.C. 75 pp.
Wagner D.L. 2005. Caterpillars of Eastern North America. Princeton University Press. Princeton, New Jersey. 256 pp.
Wagner D.L., K.J. Todd . 2016. New ecological assessment for the emerald ash borer: a cautionary tale about unvetted host-plant literature. Am. Entomol. 62:26–35.












